Abstract :
[en] Introduction. Recent availability of population-based neuroimaging datasets with extensive
psychometric characterization [1][2] opens promising perspectives to investigate the
relationships between interindividual variability in brain regions’ functional connectivity (FC)
and behavior. However, the multivariate nature of prediction models based on connectivity
severely limits interpretations from a cognitive neuroscience perspective. To address this issue,
we propose a connectivity-based psychometric prediction (CBPP) framework based on
individual region’s connectivity profile. Preliminary to the development of this region-wise
approach, we performed an extensive assessment of CBPP approaches based on whole-brain
connectivity.
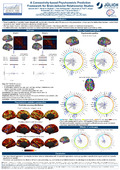
Method. Fig. 1A shows the different approaches considered at each step in our implementation
of whole-brain CBPP. We used preprocessed resting-state fMRI time series and 98
psychometric variables from the Human Connectome Project (HCP) [1]. The time series were
grouped into parcels using the Schaefer atlas [3]. FC was computed as the correlations between
the mean time series of each pair of parcels. A set of 9 confounding variables (sex, age, age2,
sex*age, sex*age2
, handedness, brain size, intracranial volume, and acquisition quarter) were
regressed out from the psychometric variables. Finally, the prediction performance for a
psychometric variable based on whole-brain FC was evaluated by the correlation between the
predicted and the true values.
For parcel-wise CBPP, the procedure was similar to whole-brain CBPP, except that the
initial features included only connectivity between a specific parcel and the rest of the brain.
We focused on the FIX-300-parcel-Pearson-SVR combination of approaches (see Fig. 1A),
which was found to be optimal for whole-brain CBPP. We illustrated two main applications to
evaluate brain region-behavior relationships in a neuroscience perspective: 1) single brain
region’s predictive power for different psychometric variables 2) variation of predictive power
across brain regions for a single psychometric variable.
Results. Fig. 1B shows the whole-brain CBPP results from all combinations of approaches, for
the 20 most well predicted psychometric variables across combinations. Overall, combinations
using FIX denoising and 200-parcel/300-parcel granularity achieved the best results, regardless
of connectivity computation method or regression method used (except multiple linear
regression).
Fig. 2A illustrates the predictive power of a pair of parcels in the Broca region for 40
selected psychometric variables. Both parcels from the left and right hemispheres exhibited
high predictive power in language-related measures, cognition composite scores and working
memory performance. Interestingly, the right parcel showed lower predictive power for
language task accuracy, but higher power for working memory abilities. These observations
are consistent with brain mapping literature about the Broca region [4][5][6][7].
Fig. 2B illustrates the variation of predictive power across parcels for working memory
task overall accuracy and face condition accuracy, as well as the HCP group activation map of
working memory task. Both predictive power spatial maps show best performing parcels in the
cingulate cortex, parietal cortex, supramarginal gyrus, lateral frontal cortex and anterior insula,
while the latter also shows well performing parcels in the inferior temporal cortex and calcarine
sulcus. High predictive power and high signal changes in group activation maps were both
found in the anterior cingulate cortex, parietal cortex, lateral frontal cortex and anterior insula.
Conclusions. We evaluated the CBPP framework using whole-brain connectivity,
demonstrating the relevance of sophisticated denoising techniques and the good performance
of simple regression-based algorithms. Based on this framework, we developed a parcel-wise
CBPP approach optimizing both prediction performance and neurobiological validity.
References
1. Van Essen, D.C., Smith, S.M., Barch, D.M., Behrens, T.E., Yacoub, E., Ugurbil, K.,
for the WU-Minn HCP Consortium (2013), 'The Wu-Minn Human Connectome
Project: An overview', NeuroImage, vol. 80, pp. 62-79
2. Caspers, S., Moebus, S., Lux, S., Pundt, N., Schütz, H., Mühleisen, T.W., Gras, V., et
al. (2014), 'Studying variability in human brain aging in a population-based German
cohort – Rationale and design of 1000BRAINS', Frontiers in Aging Neuroscience,
vol. 6, pp. 149
3. Schaefer, A., Kong, R., Gordon, E.M., Laumann, T.O., Zuo, X-N., Holmes, A.J.,
Eickhoff, S.B., Yeo, B.T.T. (2018), 'Local-global parcellation of the human cerebral
cortex from intrinsic functional connectivity MRI', Cerebral Cortex, vol. 28, pp.
3095-3114
4. Binder, J.R., Frost, J.A., Hammeke, T.A., Cox, R.W., Rao, S.M., Prieto, T. (1997),
'Human brain language areas identified by functional Magnetic Resonance Imaging',
Journal of Neuroscience, vol. 17, no. 1, pp. 353-362
5. McNealy, K., Mazziotta, J.C., Dapretto, M. (2006), 'Cracking the language code:
Neural mechanism underlying speech parsing', Journal of Neuroscience, vol. 26, no.
29, pp. 7629-7638
6. Salmon, E., Van der Linden, M., Collette, F., Delfiore, G., Maquet, P., Degueldre, C.,
Luxen, A., et al. (1996), 'Regional brain activity during working memory task', Brain,
vol. 119, no. 5, pp. 1617-1625
7. Smith, E.E., Jonides, J. (1999), 'Storage and executive processes in the frontal lobes',
Science, vol. 283, no. 5408, pp. 1657-1661